
計画研究A02 ネオ・セルフの構造的理解
▼ネオ・セルフの立体構造解析 ▼ネオ・セルフとしてのミスフォールド蛋白質解析 ▼ネオ・セルフの遺伝子解析
ネオ・セルフの立体構造解析
| 研究代表者 |  |
横山 茂之 | 理化学研究所 横山構造生物学研究室 上席研究員 理研HPへ(http://www.riken.jp/research/labs/distinguished/struct_biol/) 研究室HPへ(http://sbl.riken.jp/) |
| 分担研究者 |  |
笹月 健彦 | 九州大学 高等研究院 特別主幹教授 大学HPへ(http://ias.kyushu-u.ac.jp/organization/sasazuki_01.php) |
| 連携研究者 | 山本 健 | 久留米大学 医学部 医化学講座 教授 |
横山班 研究概要
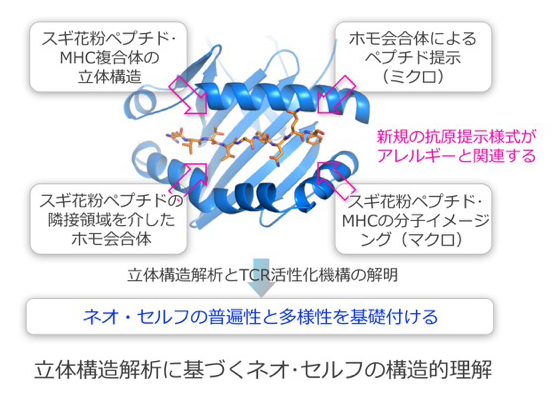
ヒトの主要組織適合性複合体(major histocompatibility complex, MHC)であるHLA分子は、自己抗原および外来抗原を特異的に結合し、細胞上でT細胞受容体(T cell receptor, TCR)に認識されます。我々は、笹月健彦(分担研究者)との共同研究により、クラスIIのHLA(HLA-II)分子であるHLA-DP5とスギ花粉抗原ペプチド(外来抗原)との複合体の立体構造解析を行い、HLA-DP5同士が会合すること、ペプチドの隣接領域によって促進されることを発見しました。本研究では、HLA-II分子と、アレルギー疾患の外来抗原、自己免疫疾患の自己抗原、がん免疫の変異抗原等との複合体(ネオ・セルフ)の立体構造解析によって、抗原提示様式を比較します。さらに、それらのネオ・セルフを認識するTCRとの3者複合体の立体構造解析によって、ネオ・セルフとTCRの結合様式を比較し、抗原提示様式や免疫応答との関係を体系化します。これらの解析により、ネオ・セルフとセルフを区別する決定的な特徴を解明し、免疫疾患の発症機構の解明や、新たな治療法の開発に貢献することをめざします。
Nature. 531(7592):122-5, 2016. doi: 10.1038/nature16991.
Nature. 520(7547):312-6, 2015. doi: 10.1038/nature14301.
J Mol Biol. 426(17):3016-27, 2014. doi: 10.1016/j.jmb.2014.06.020.
Nature. 510(7506):507-11, 2014. doi: 10.1038/nature13440.
Science. 340(6128):75-8, 2013. doi: 10.1126/science.1229521.
笹月班 研究概要
HLAは膨大な個人差(多型性)を示し、外来あるいは自己抗原由来のペプチドと特異的に結合することにより、T細胞応答を誘導し感染防御を担う一方、アレルギー疾患や自己免疫疾患を引き起こします。HLA-DP5はスギ花粉由来のペプチド(外来抗原)と結合しこれを認識するT細胞レセプターと結合し、B細胞を活性化してIgE抗体産生を誘導し、スギ花粉症(Ⅰ型アレルギー)を発症させます。同じHLA-DP5が甲状腺刺激ホルモン受容体(TSHR)由来のペプチド(自己抗原)と結合し、T細胞を活性化してB細胞にTSHRに対する自己抗体を産生させることによってグレーブス病(自己免疫疾患)が発症します。アレルギーを発症させた外来抗原ペプチドとHLAの結合様式、および自己免疫疾患を発症させた自己ペプチドとHLAの結合様式の立体構造上の差異を明らかにするため、HLA-DP5と結合するスギ花粉由来の複数のペプチドを同定し、同じくHLA-DP5と結合するTSHR由来の複数のペプチドを同定し、それぞれHLA-DP5との複合体を作製して解析を進めます。同様にグレーブス病でHLA-DR53と結合するサイログロブリン(TG)由来ペプチド、およびスギ花粉症でHLA-DR53と結合するスギ花粉由来ペプチドをそれぞれ複数同定し、それぞれHLA-DR53との複合体の高次構造を比較する。最終的にこれらと特異的に結合するT細胞受容体(TCR)とのcomplex(ペプチド・HLA-TCR complex)の高次構造の解明により、免疫システムが示す多様な免疫応答の構造学的基盤の解明を目指します。さらに制御性T細胞によるペプチド-HLA複合体の認識、骨髄移植の際の移植片対宿主(GvH)病におけるT細胞によるペプチド-HLA複合体の認識についてもペプチドの同定を目指します。また、日本人集団で著しく頻度の高いHLA-A*33:03-C*14:03-B*44:03-DRB1*13:02-DQB1*06:04-DPB1*04:01ハプロタイプがグレーブス病、橋本病、SLEさらには骨髄移植に際してのGvH病の発症を負に制御することから、このハプロタイプのどのHLAがどのようなペプチドと結合して制御するのかの解明を目指します。(これらの構造解析は横山茂之計画研究代表者との共同研究)
Blood., 125(7):1189-97, 2015. doi: 10.1182/blood-2014-10-604785.
J Clin Endocrinol Metab. 100(2):E319-24, 2015. doi: 10.1210/jc.2014-3431.
J Mol Biol. 426(17):3016-27, 2014. doi: 10.1016/j.jmb.2014.06.020.
J Clin Endocrinol Metab. 99(2):E379-83, 2014. doi: 10.1210/jc.2013-2841.
Blood. 115(23):4664-70, 2010. doi: 10.1182/blood-2009-10-251157.
N Engl J Med. 339(17):1177-85, 1998.
ネオ・セルフとしてのミスフォールド蛋白質解析
| 研究代表者 |  |
横須賀 忠 | 東京医科大学 医学部 医学科 免疫学分野 主任教授 大学HPへ(http://www.tokyo-med.ac.jp/faculty/med/course/course18.html) 研究室HPへ(http://tokyo-med-imm.jimdo.com/) |
| 分担研究者 |  |
末永 忠広 | 福島県立医科大学 医学部 准教授 |
| 連携研究者 | 荒瀬 尚 | 大阪大学 微生物病研究所 免疫学分野 教授 |
横須賀班 班研究概要
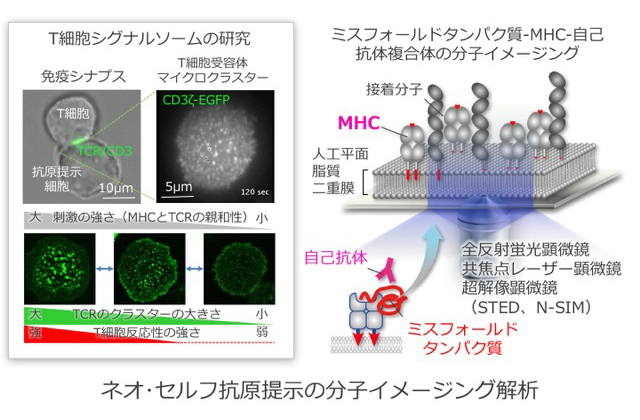
T細胞が、体内で遭遇した細胞が自分のものかどうかを識別したり、また抗原提示細胞から情報を受け取ったりする際には、二つの細胞が接着する必要があります。その接着面、いわゆる「免疫シナプス」には、数十個のT細胞受容体(T cell receptor : TCR)とMHCが集まったシグナル伝達分子の集合体「TCRマイクロクラスター」ができます。マイクロクラスターは、抗原提示細胞を模倣した人工平面脂質二重膜(プレイナーメンブレン)と1分子イメージングとの融合システムによって、研究代表者らが発見したT細胞の活性化を制御する最小シグナルユニットです。つまり、T細胞の「セルフ」の認識はTCRマイクロクラスターによってなされているのです。本研究では、これまでの膨大なT細胞シグナル研究やMHC研究の生化学・生理学的データを基盤に、先端的イメージングとの融合研究を行うことで、ミクロな視点かつ時空間的アプローチからのミスフォールド蛋白質/MHCクラスII分子複合体の構造と分子間相互作用、自己寛容破綻の誘導機構、病態解明から疾患の克服や診断技術の開発につながる分子基盤の創出を目標としています。
J Exp Med. 213(8):1609-25, 2016. doi: 10.1084/jem.20151088.
Nat Commun. 6:5555, 2015. doi: 10.1038/ncomms6555.
Nat Immunol. 15(5):465-72, 2014. doi: 10.1038/ni.2866.
Nat Immunol. 14(8):858-66, 2013. doi: 10.1038/ni.2634.
J Exp Med. 209(6):1201-17, 2012. doi: 10.1084/jem.20112741.
末永班 研究概要
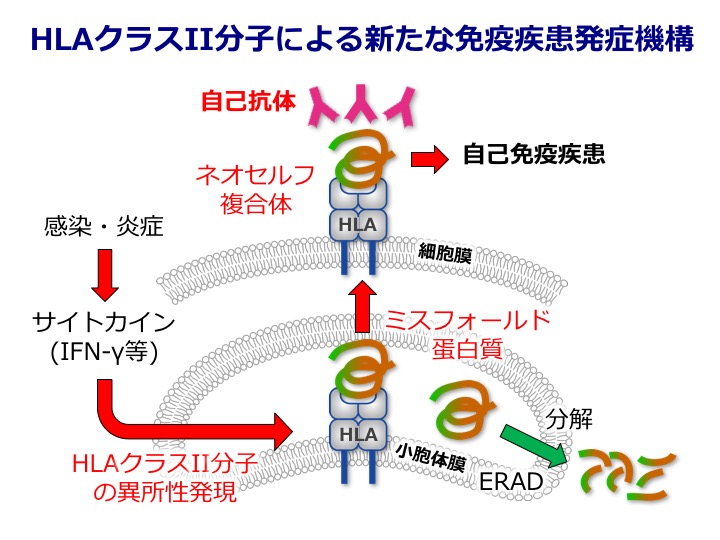
免疫システムは、通常、自己抗原に対して非応答性ですが、自己免疫疾患は自己抗原に対する免疫応答によって引き起こされると考えられております。細胞内で産生されたミスフォールド蛋白質は、通常、細胞外へ輸送されることはありませんが、それらがMHCクラスII分子と結合すると分解されずに細胞外へ輸送されることが明らかになってきました(図:左)。このようなミスフォールド蛋白質は、正常抗原とは異なる抗原性をもっていることから、ネオ・セルフ抗原として免疫疾患に関与している可能性が考えられます。実際、MHCクラスII分子によって形成されたネオ・セルフ抗原は、関節リウマチや抗リン脂質抗体症候群の患者由来の自己抗体の主要な標的抗原になっていることが明らかになってきました(図)。さらに、自己抗体のネオ・セルフ抗原に対する結合は、MHC-IIアリルによる疾患感受性と強い相関を示します。これらのことから、MHCクラスII分子によって生成されたネオ・セルフ抗原は、自己免疫疾患発症の原因になっている可能性が考えられます。本分担研究班では、MHCクラスIIによって生成されたネオ・セルフがどのように自己免疫疾患の発症に関与しているかを、TCRやBCRマイクロクラスターの1分子イメージングを用いて解析し、自己免疫疾患の発症機構を1分子レベルで明らかにしていきます。
Adv Immunol. 129:1-23, 2016. doi: 10.1016/bs.ai.2015.09.005.
Blood. 125(18):2835-44, 2015. doi: 10.1182/blood-2014-08-593624
J Biochem. 158(5):367-72, 2015. doi: 10.1093/jb/mvv093.
Proc Natl Acad Sci USA. 111(10):3787-92, 2014. doi: 10.1073/pnas.1401105111.
Int Immunol. 25(4):235-46, 2013. doi: 10.1093/intimm/dxs155.
ネオ・セルフの遺伝子解析
| 研究代表者 |  |
椎名 隆 | 東海大学 医学部 基礎医学系 分子生命科学 教授 研究室HPへ(http://mls.med.u-tokai.ac.jp/) |
| 分担研究者 |  |
細道 一善 | 金沢大学 医薬保健研究域 医学系革新ゲノム情報学分野 准教授 研究室HPへ(http://big.w3.kanazawa-u.ac.jp/) |
| 連携研究者 | 岡 晃 | 東海大学 総合医学研究所 講師 |
椎名班 研究概要
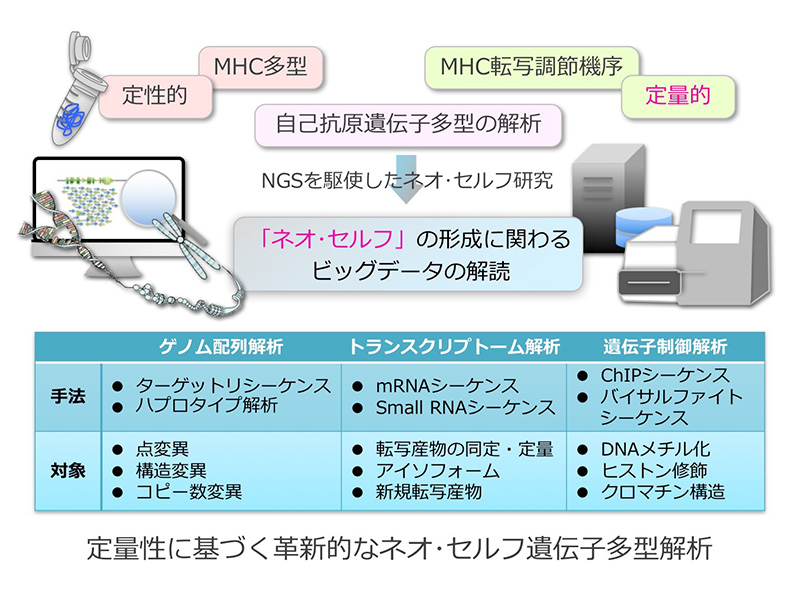
自己免疫疾患を含む100種以上の疾患が特定のHLAアレルと関連しますが、「なぜHLAと疾患が関連するのか」という、その惹起機序は長年未解決状態にあります。HLA遺伝子全領域におけるネオ・セルフ生成に関連するHLA多型や遺伝子発現における研究報告はなく、それら両面からHLA遺伝子の本質的な機能を明らかにし、ネオ・セルフ生成のメカニズムを解明する必要があります。本計画研究では、分担研究者である細道一善准教授(金沢大学医薬保健研究域医学系革新ゲノム情報学分野)とともにHLA関連疾患やアレルギーを対象に次世代シーケンサーを用いた高解像度HLAタイピング(SS-SBT)法などにより、遺伝子全領域の多型解析を実施し、健常者との関連解析から高いオッズ比や陽性率を示すHLA多型を同定します。また、解像度の高いRNA-seqにより新規のスプライスバリアントやアンチセンスRNAを検出します。さらには、メチル化解析によるHLA遺伝子発現の転写制御機構の明確化やターゲット・リシークエンシングによる転写調節因子およびその結合領域の同定も試みます。以上の最新ゲノム技術を駆使したHLA遺伝子解析により、ネオ・セルフ生成の遺伝要因の解明に挑むことを目標としています。
Immunology. 2016 Jul 10. doi: 10.1111/imm.12624.
Haematologica. 101(4):491-8, 2016. doi: 10.3324/haematol.2015.136903.
Nat Commun. 7:10356, 2016. doi: 10.1038/ncomms10356.
BMC Genomics. 16:318, 2015. doi: 10.1186/s12864-015-1514-4.
Tissue Antigens. 80(4):305-16, 2012. doi: 10.1111/j.1399-0039.2012.01941.x.
細道班 研究概要
複雑な疾患メカニズムの解明には、ゲノム配列だけでなく遺伝子を含む全ての転写産物とそれらを制御する領域ならびに因子をネットワークとして理解する必要があります。次世代シーケンサー(NGS)を始めとした最新のゲノム解析機器、一細胞レベルでの分析手法、ならびにバイオインフォマティクスを駆使した研究を通じて、ネオ・セルフ現象のメカニズムの解明に迫りたいと思います。これまで、NGSを用いたHLA領域ならびにHLA遺伝子の解析手法を開発してきましたが、その手法をさらに発展させ、HLAを中心とした免疫メカニズムの解明のためにHLA-omicsと命名した統合解析を提案しています。ゲノム、遺伝子発現、転写調節、エピジェネティクスを網羅的・統合的に解析することで、HLAに起因するネオ・セルフ現象を可視化し、そのメカニズムの解明するための手法開発を目指します。メカニズムを理解することは将来的に病態を制御することへと繋がります。すなわち、研究から得られる知見から、疾患に対する新規治療法のみならず、あらたな診断・予防法開発も期待できます。
PLoS Genet. 12(4):e1005893, 2016. doi: 10.1371/journal.pgen.1005893.
Sci Rep. 6:20611, 2016. doi: 10.1038/srep20611.
J Hum Genet. 60(11):665-73, 2015. doi: 10.1038/jhg.2015.102.
BMC Genomics. 15:645, 2014. doi: 10.1186/1471-2164-15-645.
BMC Genomics. 14:355, 2013. doi: 10.1186/1471-2164-14-355.